|
||
|
Древние Боги
|
||
 |
Развитие второй
сигнальной системы у людей ни в коем случае не было следствием разрастания
общего объема головного мозга по сравнению с объемом (весом) тела и прямо никак
не связано с этим процессом энцефализации (церебрализации) в филогении
троглодитид. Во-первых, в биологической эволюции вообще налицо тенденция
увеличения мозга (независимо от размеров тела), и по этому показателю
прямоходящие высшие приматы, включая человека, вовсе не оказываются
специфичными: степень энцефализации увеличивается у всех ископаемых
млекопитающих. По степени развития неокортекса ("новой коры"), т.е.
по степени "неоэнцефализации", можно выделить весь отряд приматов из
общей родословной млекопитающих, но невозможно отдифференцировать собственно
человеческую линию. Во-вторых, разрастание объема головного мозга в эволюции
семейства троглодитид было прямым морфологическим следствием прямохождения и
прямым биохимическим следствием плотоядения, т.е. повышенного усвоения протеина
; эти два фактора влекли за собой широкий размах индивидуальных вариаций объема
мозга, иначе говоря, делали структурно и гистологически возможными колебания в
сторону повышенной массы мозга, а естественный отбор закреплял эти отклонения,
несомненно, потому, что они были биологически выгодны. По Боне, это разрастание
мозга происходило в том же темпе, как и другие морфологические трансформации
млекопитающих в плейстоцене. Но у Homo sapiens средний размер и вес головного
мозга не возрастал и не возрастает сравнительно с поздними палеоантропами. При
этом размах индивидуальных вариаций данного признака у Homo sapiens весьма
увеличился сравнительно с палеоантропами, нередко встречается объем мозга,
значительно превышающий среднюю величину, но ничто не закрепляет этих
отклонений: они биологически нейтральны и средняя величина остается неподвижной
для черепов любого времени с верхнего палеолита до наших дней. Между тем
речевая функция мозга в корне отличает неоантропа от палеоантропа. Как видно, социальность
и разум человека никак прямо не коррелированы с тотальной величиной его
головного мозга. Зато корреляция начинает
проступать, когда измерению подвергаются по отдельности длиннотный, широтный и
высотный диаметры роста головного мозга в филогенетической цепи: шимпанзе –
австралопитек – археоантроп – палеоантроп – неоантроп. До неоантропов наиболее
интенсивно увеличивался тотальный длиннотный размер (за счет роста задних
областей); напротив, только мозг неоантропов дает интенсивный сдвиг высотного
диаметра: в лобной и теменной долях эпицентры роста перемещаются из нижних
отделов в верхние, происходит, как мы уже отмечали, усиленный рост
префронтальной области, а также поднятие и выравнивание поверхности мозга в его
своде.
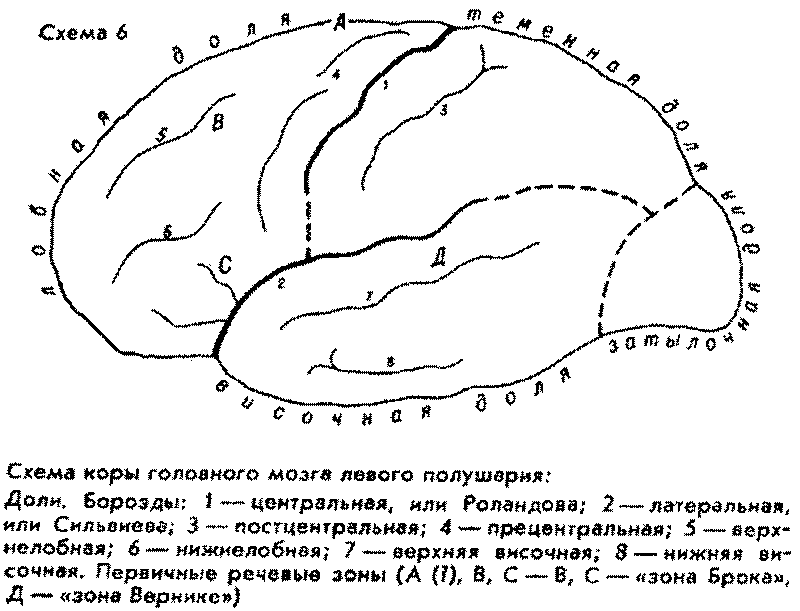
Интересно, что как раз в
самой высотной, и тем самым весьма молодой, точке свода нашего мозга
предположительно локализуется самый исходный, "инициальный" очажок
речевой функции человека (см. схему коры мозга). Правда, вопрос пока является
дискуссионным и открытым. Это так называемая "речевая зона
Пенфильда", или "верхняя речевая кора", – якобы зона начальной
речевой активизации. Эту зону Пенфильд и Роберте якобы обнаружили при
оперативных исследованиях эпилепсии в так называемом дополнительном моторном
поле на медиальной (внутренней, т.е. обращенной к другому полушарию)
поверхности в задних отделах верхней лобной извилины. Некоторые авторы считают
это прочно установленным фактом, другие при применении иных нейрохирургических
методов не обнаруживают нарушений речевых функций при поражениях этой зоны (А.
Р. Лурия) или замечают воздействие всего лишь на оттенки интонации и модуляции.
Впрочем, последнее не служило бы решающим возражением: кто знает, может быть,
так и рождалась первичная дифференциация звуков-знаков. Не будем пока ни
принимать, ни отбрасывать гипотезу Пенфильда – Робертса. Заметим лишь, что,
если бы указанная зона в самом деле все-таки оказалась начальным очажком второй
сигнальной системы, было бы поразительно, что он расположен в самой высотной
части, т.е. на самом молодом верхнем крае коры мозга Homo sapiens. В пользу
второсигнальной принадлежности этого образования (если принять описание его
Пенфильдом и Робертсом) говорит его односторонняя локализованность в
доминантном полушарии (в левом у правшей), тогда как близко расположенные
центры локализации находятся в обоих полушариях. |
|
 |
||
 |
||
 |
||